Анатомическая и экологическая специфика
видов festuca pratensis huds и festuca rupicola heuff
Мусаев
Селим Гасанпаша оглу,
доктор биологических наук, профессор,
член-корреспондент
НАН Азербайджана,
Алескеров
Мефтун Магомед оглу,
диссертант Института ботаники НАН Азербайджана.
Экологические особенности любого
растения, помимо его морфологического строения, наблюдаются также в его
анатомии. При изучении анатомического строения вегетативных и генеративных
органов в отношении разных структурных элементов, в количестве элементов и
слоев клетки, в своеобразно специализированных структурных единицах ярко
выражается его экологическая принадлежность.
К структурным элементам,
определяющим экологические особенности видов растений, относятся наличие
трихомы, толщина внешней мембраны в нижней и верхней клетке эпидермиса,
распределение устьиц в эпидермисе, степень развития механической и проводящей
тканей, количество и размер моторных клеток [Заленский 1964, Николаевский
1970]
Вид Festuca L. считается самым
распространенным видом семейства Poaceae. Представители этого вида, как сорняк,
широко распространены в малоплодородных местностях с сухим климатом, в районах
богарного земледелия, в садах, в утесистых местностях и в местностях засоления.
Кроме того, эти виды также встречаются и в горных местностях с влажным
климатом.
Анатомические и экологические
особенности видов рода Festuca неоднократно были объектом исследования многих
ученых. Приспособление растений к дефициту воды Mc Cue и Hansan (1990)
разделили на две группы: защита от засухи и стойкость к засухе. Особенности
первой группы ученые объясняют особенностями анатомического и морфологического
строения растения. К ним можно отнести наличие кутикулы и воскового слоя,
хорошо развитую склеренхиму, толщину стенки клетки, расположение устьиц своеобразной
формы и уменьшенный размер межклеточной полости [Эсау 1980, Raven 1986,
Zimmerman 1982] Eric J.V
Visser, Gerard M. Bogemann (2003), Blom V. Colmer T. D. (2000), Aulakh M. S. и
Vassmann R.(2000) в тканях некоторых видов Festuca обнаружили наличие аэренхимы
и отметили это как важный анатомический и экологический признак этих видов.
Мы в своих исследованиях, изучая
анатомическое строение двух важных представителей рода Festuca, F.pratensis
Huds. и F.rupicola Heuff, определили экологические особенности в их структуре.
Материалы
и методика исследования
Основываясь на общепринятых
методах, мы изучали продольный и поперечный срезы вегетативных органов
растений, над которыми проводились опыты. Растения были собраны в лесах Пиргулу
Шемахинского района Азербайджанской Республики в месяцах май-июнь 2007-го года.
Местность находится на высоте
Материал для анатомических
исследований был выбран из физиологически зрелых растений и зафиксирован 96%
спиртом. Срезы были проведены вручную, препараты покрашены общепринятыми методами
и помещены в глицерин-желатин. Для исследований были приготовлены постоянные и
временные препараты. При изготовлении препаратов была применена методика,
предложенная Бутник (1991) и Барыкина
(2004). Изготовленные препараты были исследованы под световыми микроскопами МБУ-3
и МБУ-6.
В создании анатомического образа
была использована терминология Захаревич (1954), Красилов (1968), Свешникова
(1970) и Гумбатов (2002).
Исследования проводились под
руководством кандидата биологических наук, доцента кафедры ботаники Гянджинской
Государственной Сельскохозяйственной Академии З. Гумбатовом.
Обсуждение
Вид Festuca pratensis – овсяница
луговая, многолетнее растение высотой 30-
Анатомическое
строение корня (рис. 1)
В поперечном срезе зрелой части
корня, которая перешла во вторичную структуру, эпиблема наблюдается как
темно-коричневая пробка - перидерма. Глубже находятся 7-8 слоев специально
расположенных паренхимных клеток. Так как во внешних 4-5 слоях клетки немного
крупноватые и расположены несколько свободно, межклеточные полости широкие.
Затем 4 слоя мелких, плотно
расположенных паренхимных клеток овальной формы, опирающихся на пояски Каспари.
Ширина межклеточной полости между паренхимными клетками обеспечивает апопластическое
движение воды, что связано с мезофитностью растения и является важнейшим
экологическим показателем.
В паренхимных клетках,
расположенных ближе к центру, встречаются запасные питательные вещества.
Последний слой паренхимы состоит из более плотно расположенных мелких клеток и
опоясывает эндодерму. Во внешнем слое центрального цилиндра видна однослойная
оболочка, состоящая из клеток Каспара пятиугольной формы. Внешний слой этих
клеток изнутри с трех сторон имеет утолщения, из-за чего под микроскопом
наблюдается в форме подковы [Гумбатов 2002, стр.72].
На поперечном срезе корня видно,
что паренхимный слой занимает больше места, чем центральный цилиндр.
Центральный цилиндр заполняют
первичные элементы ксилемы и флоэмы. Между мелкими элементами наблюдается 6
крупных метаксилемных сосудов. Ближе к периферии расположено 14 протоксилемных
сосудов и рядом с ними находятся элементы протофлоемы.
На продольном срезе корня видны
пористые утолщения трахеид.
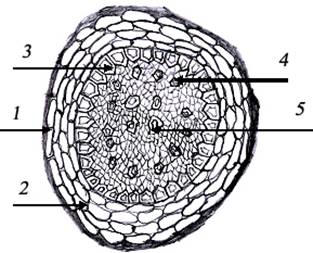
Рис 1. Анатомическое
строение корня: 1) перидерм; 2) паренхима; 3) пояска Каспари; 4) протоксилемные
сосуды; 5) метаксилемные сосуды.
Анатомическое
строение стебля (рис. 2)
Стебель Festuca pratensis тоже
имеет типичное мезофитное строение. На поперечном срезе стебля видно, что он с
внешней стороны покрыт однослойным эпидермисом. Глубже от эпидермиса расположен
слой механической ткани, состоящий из волокон склеренхимы. Между волокнами
имеются еще не завершившие дифференциацию мелкие проводящие пучки. Особо важной
биологической спецификой стебля является наличие множества проводящих пучков.
Дальше от слоя склеренхимы между паренхимными клетками расположено 14-15 зрелых
проводящих пучков замкнуто-коллатерального типа. Они обладают склеренхимным и
паренхимным покровом. Такое расположение механической ткани и проводящих пучков
придают стеблю устойчивость.
В пучках ближе к перидерме
расположена флоэма, а ближе к центру - 3-4 метаксилемных и множество мелких
ксилемных сосудов.
По классификации Р.М.Середина
мелкие проводящие пучки, дифференциация которых завершена, делятся на два типа:
I - окруженные склеренхимой со всех сторон и II - только с трех сторон
окруженные склеренхимой. Зрелые проводящие пучки только одной стороной
опираются на склеренхиму и относятся III типу.
На продольном срезе стебля с
внешней стороны наблюдается наличие кутикулы. В паренхиме клетки крупные и
межклеточные полости широкие. Эти клетки, также называемые клетками Стразбургер,
играют вспомогательную роль при транспорте органических веществ к флоэмным сосудам.
Во внутренних слоях ксилемных сосудов тоже встречаются пористые и спиральные
утолщения.
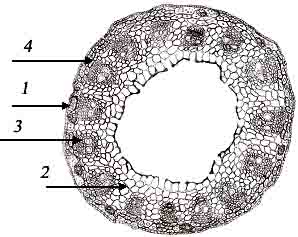
Рис. 2.
Анатомическое строение стебля: 1) эпидермис; 2) паренхима; 3) проводящие пучки;
4) склеренхима.
Анатомическое строение листа (рис. 3)
Многие жизненно важные для
растений физиологические процессы протекают в листьях. Поэтому анатомическая
структура листа богата интересными элементами строения. В разных видах эти
элементы были исследованы Цвелев, Сергеева,
Мусаев, (1982), Прокудин (1977) и другими учеными.
На поперечном срезе лист согнут в
виде «V». Эпидермис покрыт кутикулой. На поверхности эпидермиса волоски не
встречаются. И в нижнем и в верхнем эпидермисе имеются устьица, что является важным
биологическим показателем. На верхнем эпидермисе наравне с проводящими пучками
расположены моторные клетки. У этих видов моторные клетки, расположенные вокруг
центральной жилки, хорошо развиты и более крупные. Это связано с мезофитностью
этого вида, чем он и отличается от ксерофитного сорта.
Мезофилла листа образована
проводящими пучками и склеренхимной тканью. Количество проводящих пучков может
быть разным, по мнению некоторых специалистов оно может достигать 27 [Мусаев, Сергеева, 1982].
Проводящие пучки листа с обеих
сторон прикреплены склеренхимой к эпидермису. Такое строение, по классификации
Вуколова, относится к 5-ому типу. Среди зрелых пучков 5-ого типа видны мелкие
пучки 6-ого типа, не имеющие фиксации склеренхимой в отношении 1:1. В части
ксилемы, где пучки направлены к верхней части эпидермиса, расположено 3-4
крупных метаксилемных сосудов. Проводящие пучки окружены паренхимными клетками.
Это свойственно злакам, эти клетки богаты такими органеллами как митохондрии,
микротела и хлоропласты. Благодаря наличию таких клеток фотосинтез протекает
путем С-4 [Эзау 1980, стр.368]. Этот путь фотосинтеза, наряду со злаками, еще
встречается у 10 семейств. Эта особенность зерна дает возможность в разных
экологических условиях более эффективно используя углерод стать доминирующим
растением.
На продольном срезе листа в
центральной жилке видны крупные кольцеобразные трахеиды и сопровождающие ее два
пористых сосуда, несколько ситовидных трубок и сопровождающие их спутниковые
клетки. В боковых проводящих пучках встречаются спиральные сосуды. Паренхима,
формирующая мезофилл листа, состоит из крупных, свободно расположенных клеток.
Если посмотреть на продольный срез эпидермиса, можно увидеть устьица,
расположенные в шахматном порядке, а в нижнем эпидермисе они расположены по-прямой.
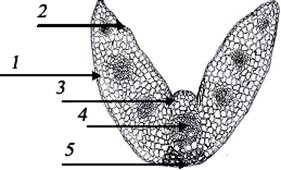
Рис. 3.
Анатомическое строение листа: 1) эпидермис; 2) устицы; 3) моторные клетки; 4)
проводящие пучки; 5) склеренхима.
Анатомическое строение корня (рис. 4)
Другой объект исследования, вид
Festuca rupicola Heuff, многолетнее растение темно-зеленого цвета, образует густой
дерн. Имеет стебель высотой 30-
В препаратах, изготовленных из
зрелой части корня, на поперечном срезе видно, что с внешней стороны она
покрыта пробковой корой. Внутри расположены паренхимные клетки, образующие 3-4
ряда. В отличие от предыдущего рассматриваемого вида, здесь большую часть
клетки занимает центральный цилиндр, а ткани паренхимы наблюдаются как внешний
слой. Два внешних слоя паренхимы состоят из свободно расположенных крупных
клеток, а два внутренних слоя - из плотно расположенных овальных клеток.
Центральный цилиндр, окруженный
клетками Каспара, окружен элементами флоэмы и ксилемы. Количество проводящих
сосудов, по сравнению с Festuca pratensis, мало. Так как в центральной части расположено
4 крупных метаксилемных сосудов, а ближе к внешней поверхности - 12 протоксилемных
сосудов, протофлоэмные клетки граничат с элементами протоксилемы.
На продольном срезе корня
наблюдаются пористые и кольцевые утолщения элементов ксилемы.
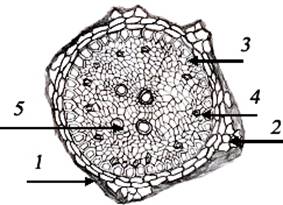
Рис. 4. Анатомическое
строение корня: 1) перидерм; 2) паренхима; 3) пояска Каспари; 4) протоксилемные
сосуды; 5) метаксилемные сосуды.
Анатомическое строение стебля (рис. 5)
Стебель с внешней стороны покрыт
однослойным эпидермисом, обладающим толстой кутикулой. Клетки эпидермиса
мелкие, ровные, овальной формы. Волоски на эпидермисе встречаются редко.
Волокна склеренхимы под эпидермисом расположены в 3-4 слоя и образуют плотную
опору в виде окружности. Виду F. Rupiсola присуща хорошо развитая механическая
ткань, что является одной из особенностей ксерофитизма.
Глубже от механической ткани
расположены клетки паренхимы. По направлению к центральной полости размер
клетки паренхимы и межклеточные полости увеличиваются. Наравне с проводящими пучками
склеренхима, образуя несколько дополнительных слоев, сливается с ними. Таким
образом, по классификации Р.М.Середина и М.Галкина проводящие пучки стебля
относятся к 3-му типу. Количество элементов и проводящих пучков F.rupicola по
сравнению с прежним, мезофитным виду, намного меньше, что и считается важнейшим
биоэкологическим показателем. 7 замкнуто-коллатеральных проводящих пучков,
расположенных в стебле F.rupicola, с внешней части окружены покровами склеренхимы
и паренхимы. Элементы метаксилемы расположены по три. На продольном срезе под
эпидермисом наблюдаются хорошо развитые волокна склеренхимы, пористые и
кольцевые утолщения ксилемных сосудов. Структура листьев более богата
признаками ксерофитизма.
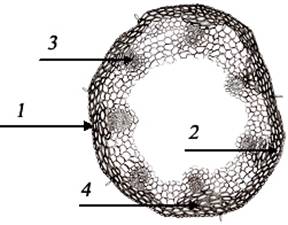
Рис. 5.
Анатомическое строение стебля: 1) эпидермис; 2)паренхима; 3) проводящие пучки;
4) склеренхима.
Анатомическое строение листа (рис. 6)
Клетки эпидермиса, покрывающие
листья вида F.rupicola, на верхней стороне мелкие, а на нижней - немного
крупнее и вытянуты в длину. Над поверхностью эпидермиса видна толстая кутикула,
волоски расположены редко по три. Устьица имеются только на верхнем эпидермисе.
Волокна склеренхимы расположены в нижнем эпидермисе, под центральными проводящими
пучками и еще в виде нескольких слоев на границе с верхним эпидермисом.
На поперечном срезе лист
F.rupicola свернут, образуя окружность, что и показывает наличие моторных
клеток, расположенных в особом порядке. Здесь же находится 5-6 моторных клеток
вдоль борозды, разделяющей 5-6 жилок. При дефиците воды эти клетки переходят в
состояние плазмолиза, что заставляет лист согнутся внутрь, в результате чего
устьица, расположенные на поверхности эпидермиса, замыкаются и препятствуют
потере воды. В это же время уменьшается поверхность листа, и нижний слой
эпидермиса, покрытый кутикулой, направлен в сторону солнца. Клетки паренхимы
крупные и хорошо развитые. Между этими клетками расположено 7 проводящих
пучков. Мелкие пучки, расположенные в центре и вдоль ребра, относятся к 5-у
типу, а крайние пучки 6-ого типа ни с какой стороны не соприкасаются со
склеренхимой.
В центральных пучках расположено
4 крупных метаксилемных сосудов, в боковых пучках эти сосуды меньше размером и
их всего 2-3.
На продольном срезе в центральной
жилке видна одна крупная трахеида, имеющая кольцеобразное утолщение, и
несколько мелких трахеид, имеющих пористые утолщения. Паренхимные клетки плотно
расположены и межклеточная полость меньше.
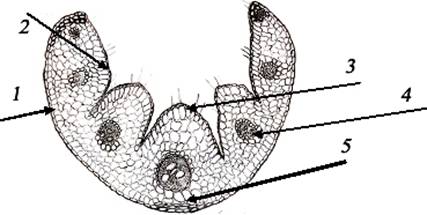
Рис. 6.
Анатомическое строение листа: 1) эпидермис; 2) устицы; 3) моторные клетки; 4)
проводящие пучки; 5) склеренхима.
Результаты
В анатомическом строении вида
Festuca pratensis Huds были обнаружены элементы, связанные с метофитизмом:
- на поперечном срезе корня
видно, что паренхима занимает относительно большую площадь, чем центральный
цилиндр;
- в листьях, в стебле и в корне
межклеточные полости, обеспечивающие апопластическое движение воды между
клетками, широкие;
- количество проводящих сосудов и
пучков в листьях, в стебле и в корне намного больше;
- лист плоский, при сгибе
замыкается не полностью, из-за наличия моторных клеток рядом с центральной
жилкой, принимает «V» -образную форму;
- на эпидермисе волоски не
встречаются;
- устьица расположены и в нижнем
и в верхнем эпидермисе листьев.
Признаки ксерофитизма вида
Festuca rupicola Heuff выражаются в нижеприведенных свойствах:
- основную часть корня занимает
центральный цилиндр;
- полости между паренхимными
клетками маленькие;
- корень, стебель и листья
покрыты толстой кутикулой;
- число проводящих пучков меньше;
- ткани склеренхимы сильно
развиты;
- на поверхности листьев и стебля
имеются волоски;
- устьица расположены только на
верхнем эпидермисе листа;
- лист согнут по окружности, в
результате чего поверхность верхнего эпидермиса, где расположены устьица,
замыкается.
Литература
1.
Бутник А.А. и др. Экологическая анатомия пустырных
растений Средней Азии. Ташкент, 1991.
2.
Гумбатов З.И. Морфология и анатомия растений.
Гянджа, 2002.
3.
Галкин М.А, Середин Р.М. О микроструктуре листьев
мятликов флоры Северного Кавказа // Биолог. Науки, 2, 1973.
4.
Заленский В.В. Материалы количественной анатомии
различных листьев одних и тех же растений. Киев, 1904
5.
Красилов В.А. Электронная микроскопия замыкающих
клеток устьиц // Палеонтолог. журнал, 3, 1978, стр.128-130.
6.
Мусаев С.Г, Сергеева В.В. Морфолого-анатомический
анализ листьев и стеблей некоторых видов Овсяниц (Festuca L.) флоры Северного
Кавказа и восточного Азербайджана. Баку, 1982.
7.
Николаевский Б.Г. К экологии анатомии листа видов
рода Festuca.L. // Украинский
бот. журнал, 1970, а. 27; 5.
8.
Прокудин Ю.Н. Злаки Украины. Киев, Наукова Думка,
1977.
9.
Барыкина Р.П., Василова Т.Д., Справочник по
ботанической микротехнике. М., Изд. Московского университета, 2004.
10.
Свешникова И.Н. О терминологии эпидермы листа
современных и ископаемых хвойных // Ботанический журнал, 1970, 4, с. 510-524.
11.
Сергеева
В.В. Род Festuca. L во флоре Северного
Кавказа, Краснодар, 1982.
12.
Середин Р.М.
Дикорастующие злаки Северного Кавказа: Дисс. ... докт. биол. наук.- Пермь,
1967.
13.
Цвелев Н.Н.,
Злаки СССР. Наука, Ленинград, 1976.
14.
Эзау К.
Анатомия семенных растений. 1-2 том, М., Мир, 1980.
15.
Aulakh M.S, Wassmann R, Rennenberg H and Fink S, 200. Pattern and
amount of aerenchyma relate to variable methane transport capacity of different
rice cultivars. Plant Biology, 2, 182-194.
16.
Eric J.W. Visser and Gerard M. Bogemann. Measurement
of porosity in very small samples of plant tissue. Plant and soil 253. 81-90.
2003,
17.
Mc Cue, K.F., Hanson, A.D., 1990. Drought and
salt tolerance: towards understanding and application. Trends in biotechnology.
8: 358-362.
18.
Raven P.H, Evert, R.F., Eichorn, S.E. 1986:
Biology of plants, 4th edition,
19.
Visser EJW, ColmerTD, Blom CWPM and Vosenek
LACJ, 2000. Changes in growth, porosity and radial oxygen loss from
adventitions roots of selected mono and dicotyledonous wetland species with
contrasting types of aerenchyma. Plant Cell Environ, 23, 1237-1245.
20.
Zimmermann, M.H.,
Поступила в редакцию 04.05.2009 г.