Явление саморегуляции двигательной единицы
Васильев Геннадий Федорович.
Вводная часть
Настоящая работа относится к области физиологии мышц в биологии и объясняет процесс регуляции сокращения скелетных мышц.
Известно, что скелетная мышца состоит из двигательных единиц, представляющих собой группу мышечных волокон, иннервируемых одним двигательным нейроном, и, поэтому, являющихся минимальной структурно-функциональной единицей двигательной системы.
По современным представлениям двигательная единица состоит из экстрафузальных мышечных волокон (далее, ЭМВ), производящих работу при сокращении (эффектор), и мышечного веретена (fusus – веретено), являющегося группой особых мышечных волокон с рецепторами растяжения в их центральной части. Эти мышечные волокна заключены в соединительнотканную капсулу и поэтому названы интрафузальными мышечными волокнами (далее, ИМВ). ИМВ расположены параллельно ЭМВ и прикреплены своими концами к концам ЭМВ так, что при изменении длины ЭМВ, в той же степени изменяется и общая длина ИМВ. Важно заметить, что мощность ЭМВ существенно выше мощности ИМВ, поэтому при взаимодействии ЭМВ и ИМВ длина двигательной единицы всегда определяется длиной ЭМВ, а длина ИМВ вынуждена отслеживать длину ЭМВ. Наличие соединительнотканной капсулы обеспечивает благодаря хорошему скольжению полную механическую изоляцию частей ИМВ, заключенных между их концами, от воздействия на них ЭМВ. Схематично двигательная единица представлена на рис. 1.
| Смотреть ГДЗ по геометрии 9 класс Атанасян смотреть www.euroki.org |
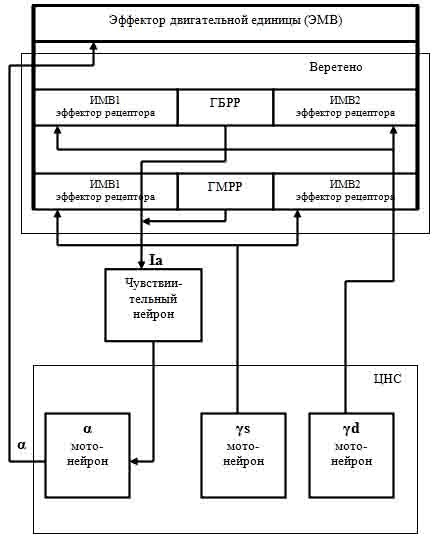
Рис. 1. Схема двигательной единицы.
радиаторы по доступным ценам от бренда WWQ stron-parts.ru
Эфферентная иннервация мышечных волокон осуществляется из центральной нервной системы (далее, ЦНС) через аксоны мотонейронов, расположенных в спинном мозге. ЭМВ управляется через аксон альфа-мотонейрона, а ИМВ – через аксоны гамма-мотонейронов (их – два, и подробнее об этом будет сказано далее). Поскольку каждое мышечное волокно ИМВ вследствие нахождения в его центральной части рецептора растяжения разделено на две части (на рис. 1: ИМВ1 и ИМВ2), аксон гамма-мотонейрона, иннервирующий его, на конце раздваиваются и обеспечивают эфферентную иннервацию каждой из этих двух частей. Таким образом, возбуждение гамма-мотонейрона приводит к одновременному сокращению двух частей мышечного волокна ИМВ, что создает стимул для рецептора растяжения. Исследования показали, что рецепторы растяжения с высокой чувствительностью реагируют на амплитуду и скорость растягивающего воздействия.
Как замечено ранее, в двигательной единице имеется два гамма-мотонейрона (обычно их обозначают, как γs-мотонейрон и γd-мотонейрон), и каждый из них возбуждает свои ИМВ. Благодаря этому обстоятельству, имеющиеся в мышечном веретене волокна, образуют две группы ИМВ. В каждую группу может входить различное количество ИМВ (от 1 до 10). Эти две группы волокон различаются по быстродействию. Многие авторы публикаций рецепторы группы ИМВ, возбуждаемой γs-мотонейроном, называют медленными рецепторами (на рис. 1 обозначены, как ГМРР), а рецепторы группы ИМВ, возбуждаемой γd-мотонейроном, - быстрыми (на рис. 1 обозначены, как ГБРР).
С каждым рецептором растяжения любого ИМВ контактируют и воспринимают сигнал возбуждения рецептора аннулоспиральные окончания разветвления афферентного нервного волокна, являющегося дендритом чувствительного нейрона двигательной единицы. Благодаря этому, чувствительный нейрон суммирует импульсы возбуждения от всех рецепторов мышечного веретена (суммарный сигнал обычно обозначается – Ia). Аксон чувствительного нейрона подходит к телу альфа-мотонейрона двигательной единицы и обеспечивает его возбуждение при растяжении рецепторов ИМВ. Аксон альфа-мотонейрона разветвляется на конце и возбуждает каждое мышечное волокно ЭМВ двигательной единицы. Сигнал, возбуждающий ЭМВ в литературе обычно обозначается, как α.
В ходе различных экспериментов было замечено, что, если принудительно сжать ЭМВ вдоль оси, то ИМВ теряют натяжение и сигнал на их выходе пропадает. Этого не происходит при изменении естественных состояний мышцы, и, следовательно, существует оперативная связь, обеспечивающая соответствие между собой длины ЭМВ и ИМВ. Попытки проследить эту связь не дали результата. В последствии явление адекватного возбуждения альфа- и гамма-мотонейронов получило название «коактивация альфа- и гамма-мотонейронов при движениях» или «альфа-гамма-сопряжение».
Valbo A. B. (1971), [3] установил, что при естественном сокращении мышцы (в эксперименте сгибался палец испытуемого), импульсация афферентов мышечных веретен (сигнал от рецепторов ИМВ) возникает с короткой задержкой после начала сокращения, что противоречило гипотезе коактивации. Осциллограмма этого эксперимента приведена в [2] (с. 97, рис. 5.9). Однако из этого и других экспериментов делается вывод, что альфа- и гамма-мотонейроны возбуждаются все-таки одновременно, но импульсация афферентов мышечных веретен отстает из-за низкой скорости проведения по гамма-волокнам и латентного периода сокращения интрафузальных (мышечных) волокон. Такое объяснение поддерживало гипотезу коактивации. Согласившись с этим, Р. Шмидт и М. Визендангер (1996) предполагают (см. [2], с. 97, колонка 1, абзац 3), что (цитирую) «главное назначение гамма-иннервации, возможно, в том, чтобы предотвратить во время сокращения экстрафузальных волокон расслабление мышечных веретен, а, следовательно, сохранить их адекватную рецепторную функцию» (конец цитаты).
Таким образом, гипотеза альфа- гамма-сопряжения предполагает, что в процессе биорегуляции высшие отделы ЦНС на основании сигналов от рецепторов вырабатывают комплексный сигнал управления двигательной единицей, который поступает из высших отделов ЦНС по двум сопрягаемым каналам: каналу альфа-мотонейрона и каналам гамма-мотонейронов. При этом по каналу альфа-мотонейрона ЦНС управляет сокращением двигательной единицы. Но, т.к. при сокращении двигательной единицы рецепторы растяжения теряют натяжение и, следовательно, способность воспринимать информацию, ЦНС по каналам гамма-мотонейронов восстанавливает натяжение, и способность адекватно воспринимать информацию восстанавливается.
Такая интерпретация назначения сигналов управления предполагает, что ЦНС непосредственно управляет сокращением ЭМВ (используя информацию от рецепторов растяжения ИМВ Ia, и формируя сигнал управления α) и, следовательно, включена в замкнутый контур регуляции.
Однако, гипотеза альфа- гамма-сопряжения не получила развития из-за своей непродуктивности. Видимо, по причине отсутствия продуктивных гипотез, в последних отечественных публикациях (см., например, [1]) какие-либо гипотезы по организации и функционированию системы управления движением вообще не упоминаются. Авторы приводят только описания элементов системы. Это свидетельствует о стагнации. Остановка в развитии свидетельствует об ошибках в интерпретации явления, потому что правильная интерпретация всегда эффективна и всегда открывает перспективы развития.
Основная часть
Проведенные исследования позволяют утверждать, что высшие отделы ЦНС не вовлечены в замкнутый контур биорегуляции двигательной единицы и не формируют сигнал управления, действующий в контуре регуляции. Высшие отделы ЦНС формируют сигналы задания, содержащие информацию о необходимых параметрах движения, и поэтому находятся вне замкнутого контура регуляции. При этом сигналы задания по положению, используемые в режиме слежения, от ЦНС поступают на гамма-мотонейроны, а на альфа-мотонейрон из центра рефлексов ЦНС поступает не сигнал задания по положению, а сигнал задания по скорости и ускорению, используемый для повышения качественных характеристик движения (преодоление инерции масс и сил сопротивления).
Структурная организация двигательной единицы позволяет ей самостоятельно осуществлять отработку поступающих из ЦНС сигналов задания по положению (на гамма-мотонейроны) и сигналов задания по скорости (на альфа-мотонейрон), что повышает качество движения.
Таким образом, связь между сокращением ИМВ и ЭМВ не является следствием сопряженного управления из ЦНС. Сокращение ЭМВ адекватно сокращению ИМВ в результате причинно-следственной связи, возникающей в результате работы системы саморегуляции двигательной единицы.
Основным доказательством достоверности данного утверждения являются результаты системного анализа структуры двигательной единицы.
Итак (см. рис. 1), каждое ИМВ содержит в средней своей части рецептор растяжения, и, в сущности, представляет собой два последовательно соединенных мышечных волокна, разделенных между собой рецептором растяжения. Оба указанных мышечных волокна иннервируются одним и тем же аксоном гамма-мотонейрона, благодаря его разветвлению. Таким образом, возбуждение гамма-нейрона приводит к сокращению двух синхронных антагонистов, что, в свою очередь, приводит к увеличению стимула для рецептора растяжения.
ЭМВ является эффектором системы, а степень его сокращения является регулируемой величиной (далее, РВ). Поскольку мощность ЭМВ намного превосходит мощность всех ИМВ, то общая длина каждого ИМВ вместе с длиной рецептора всегда вынуждена соответствовать длине ЭМВ. При такой встройке рецептора растяжения, сокращение ИМВ его растягивает, а сокращение ЭМВ – уменьшает растяжение. Таким образом, результирующим стимулом для рецептора растяжения является разность между сокращением ИМВ и сокращением ЭМВ. Системный подход позволяет интерпретировать это устройство, как устройство сравнения. При этом сокращение ИМВ интерпретируется, как сигнал задания (далее, СЗ), потому что в результате сокращения ИМВ увеличивается растяжение рецептора и, следовательно, сильнее сокращается ЭМВ. Сокращение ЭМВ, уменьшающее растяжение рецептора, интерпретируется, как сигнал отрицательной обратной связи (далее, СОС), потому что величина сокращения ЭМВ это - РВ. Результатом работы этого узла сравнения является стимул рецептора растяжения. Следовательно, результирующий стимул рецептора растяжения является сигналом управления (далее, СУ) для системы саморегуляции.
Сокращение ИМВ обеспечивают через гамма-мотонейрон высшие отделы ЦНС, поэтому высшие отделы ЦНС интерпретируются, как система верхнего уровня, формирующая сигнал задания по положению для системы саморегуляции.
Каждый рецептор растяжения ИМВ охвачен аннулоспиральным окончанием разветвления афферентного нервного волокна, являющегося дендритом чувствительного нейрона. Это обеспечивает суммирование в чувствительном нейроне всех сигналов от всех рецепторов. Аксон чувствительного нейрона подходит к телу альфа-мотонейрона, что обеспечивает доставку к нему суммарного СУ. В свою очередь, аксон альфа-мотонейрона иннервирует все ЭМВ двигательной единицы, что и обеспечивает замыкание контура системы управления по положению.
Последовательная цепочка: рецептор растяжения – чувствительный нейрон – альфа-мотонейрон – ЭМВ интерпретируется, как исполнительный механизм (далее, ИМ) системы саморегуляции.
Описанная структура иллюстрируется функциональной схемой двигательной единицы (см. рис. 2).
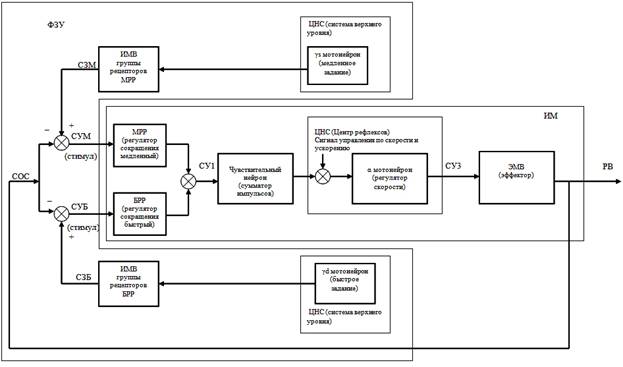
Рис. 2. Функциональная схема двигательной единицы.
Совместная работа указанных выше элементов в системе при любом текущем значении СЗ (при любой степени сокращения ИМВ) сводится к поддержанию ИМ такого текущего значения РВ (степень сокращения ЭМВ), при котором формирователем закона управления (ФЗУ) вырабатывается текущий СУ (величина стимула для рецептора растяжения), необходимый ИМ, для поддержания именно этого текущего значения РВ. Это означает, что имеет место следящая система и, что при управлении по положению «провисания» ИМВ не допускают в процессе регуляции сами ЭМВ.
Возникший в описанном процессе сигнал управления обеспечивает лишь достижение заданного положения. Но он не содержит информационных ресурсов, необходимых для обеспечения требуемой скорости и ускорения перемещаемых масс. Системы, не содержащие таких информационных ресурсов в сигнале управления, отрабатывают задание по положению не эффективно (медленно разгоняются, имеют большое отставание от задания при движении в следящем режиме).
Известны методы повышения эффективности работы систем управления по положению. Один из них заключается в добавлении в систему сигнала управления по скорости. Система управления двигательной единицей поддерживает именно этот метод повышения качества управления. Сигнал управления по скорости вводится из высших отделов ЦНС на вход альфа-мотонейрона. Этот сигнал управления формирует система рефлексов (безусловных и условных), которая обеспечивает качество движения и при управлении, и в условиях возмущения по нагрузке (динамические силы, силы пассивного и активного сопротивления).
Конечно, сигнал управления по скорости, подаваемый на альфа-мотонейрон, будет конкурировать с сигналом управления по положению, подаваемым на гамма-мотонейрон. Это хорошо видно в результатах эксперимента упомянутого в водной части A. B. Valbo.
Достоинством этого эксперимента A. B. Valbo было то, что палец сгибался не один, а три раза подряд. Сравнивая сегодня между собой эти три осциллограммы [2] (с. 97, рис. 5.9), можно увидеть, что при первом сгибании отставание было примерно – 0,25 с, при втором, примерно – 0,35 с., при третьем, примерно – 0,5 с. Т.е. отставание нарастало, и его невозможно объяснить свойствами элементов структуры, как это было тогда сделано. Но эти результаты эксперимента вполне объяснимы рефлексом форсировки, поскольку он запускается непосредственно через альфа-мотонейрон (контур управления по скорости). За счет рефлекса форсировки, ЭМВ сокращаются несколько быстрее, чем того требует сокращение ИМВ. В результате, растяжение рецепторов ИМВ под действием сигнала задания из головного мозга несколько задерживается, и сигнал управления по положению на время разгона уменьшается или пропадает полностью. Нарастание отставания в последующих сгибаниях пальца объясняется тем, что система при каждом сокращении тренировалась, и эффективность форсировки повышалась.
Ранее было замечено, что высшие отделы ЦНС для управления по положению подают на двигательную единицу два сигнала задания, поступающие, соответственно, от γs-мотонейрона на медленные рецепторы и от γd-мотонейрона на быстрые рецепторы. Известно, что такое разделение обеспечивает раздельное управление позой и активным движением. Описанная структура (см. рис. 2) обеспечивает параллельную отработку обоих заданий.
Область научного и практического значения
Исследованиями двигательной системы занимались многие талантливые ученые, и была проделана огромная работа. Но великолепными результатами исследований можно будет воспользоваться в полной мере только после описания работы двигательной системы в целом. Назрела необходимость системного подхода, раскрывающего, где возникает информация управления, как она преобразуется и в каком направлении передается. Поэтому данная научная работа является актуальной.
Результаты работы позволяют уточнить знания в биологии, повысить качество диагностики и лечения в медицине, а также качество подготовки специалистов в практических областях деятельности. Все это открывает широкие перспективы развития физиологии мышц, в т.ч. с использованием кибернетических методов.
Литература
1. Орлов Р. С., Нормальная физиология: учебник, 2-е изд., испр. и доп. – М. : ГЭОТАР – Медиа, 2010. – 832 с. : ил., с. 221 – 226.
2. Р. Шмидт, М. Визендангер, Физиология человека: в 3-х томах: перевод с англ, под ред. Р. Шмидта и Г. Тевса (Мир, 1996), с. 88-128.
3. Valbo A. B. (1971). Muscle spindle response at the onset of isometric voluntary contractions in man. Time difference between fusimotor and skeletomotor affects. J. Physiol. (Lond.), 218, 405.
Поступила в редакцию 22.03.2013 г.